Le programme de génétique, en terminale spécialité SVT, suscite régulièrement des difficultés aux élèves. Afin de proposer une remédiation et une aide aux élèves, j’ai réalisé une activité reliant des observations réelles et virtuelles. Grâce au logiciel en ligne Mesurim2 de Philippe Cosentino, il est possible d’exploiter des modèles 3D, afin d’appréhender, grâce à l’étude statistique, les notions de base de la génétique, tout en s’appuyant sur des éléments concrets, tout en étant en classe entière ou à la maison. Nous avons travaillé sur les lois de Mendel et l’approfondissement des notions de brassages inter et intrachromosomique sur un modèle différents des drosophiles observées en TP, les grains de maïs.
Professeur expérimentateur
| Amaury TAVERNIER, au lycée Jeanne d’Albret à Saint Germain-en-Laye (78) |
Vidéo de présentation
Caractéristiques de la séquence
| LIAISON AVEC LE PROGRAMME | |
|---|---|
| Niveau concerné | Terminale spé SVT |
| Partie du programme | Génétique et évolution, L’origine du génotype des individus |
| PLACE DANS LA PROGRESSION |
|---|
| Lors du chapitre sur l’origine du génotype des individus, les activités ont été proposées pour illustrer les lois de Mendel, puis suite au TP « classique » sur les brassages, réalisé avec l’observation des drosophiles, cette activité a été l’objet d’un approfondissement et retour sur les apprentissages en classe entière et en travail à la maison. |
| MOTIVATION |
|---|
|
| PROBLÈME À RÉSOUDRE |
|---|
Comment interpréter les résultats de croisements avec transmission d’une ou deux paires d’allèles (liés ou non entre eux) ? |
| NOTIONS, SAVOIR-FAIRE, compétences | |
|---|---|
| Notions | Extrait du BO spécial n° 8 du 25 juillet 2019 : Le brassage des génomes à chaque génération : La reproduction sexuée des eucaryotes : La fécondation entre gamètes haploïdes rassemble, dans une même cellule diploïde, deux génomes d’origine indépendante apportant chacun un lot d’allèles. Chaque paire d’allèles résultant est constituée de deux allèles identiques (homozygotie) ou de deux allèles différents (hétérozygotie). En fin de méiose, chaque cellule produite reçoit un seul des deux allèles de chaque paire avec une probabilité équivalente. Pour deux paires d’allèles, quatre combinaisons d’allèles sont possibles, équiprobables ou non en cas de gènes liés. Le nombre de combinaisons génétiques possibles dans les gamètes est d’autant plus élevé que le nombre de gènes à l’état hétérozygote est plus grand chez les parents. Comprendre les résultats de la reproduction sexuée : Principes de base de la génétique. L’analyse génétique peut se fonder sur l’étude de la transmission héréditaire des caractères observables (phénotype) dans des croisements issus le plus souvent de lignées pures (homozygotes) et ne différant que par un nombre limité de caractères. |
| Compétences ou Capacités expérimentales |
|
| Compétences transversales | Mise en relation réel et virtuel |
| Cadre de référence des compétences numériques (CRCN) | |
|---|---|

|
|

|
|

|
|

|
|
Déroulement de la séquence
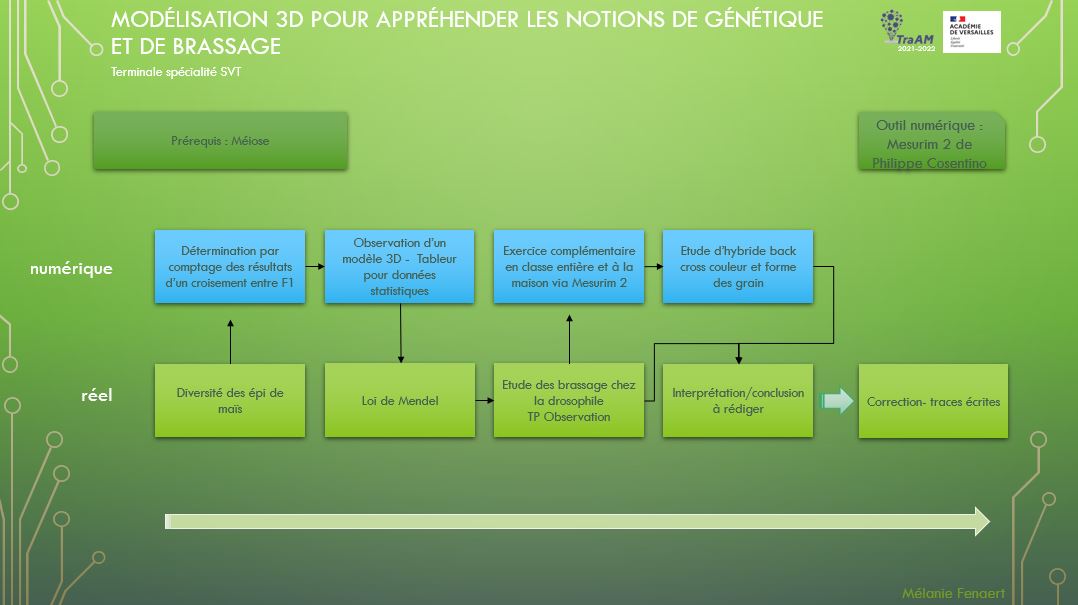
| ACTIVITÉ | ||
|---|---|---|
| Durée :
– Activité 1 : 0h30 – Activité 2 : 1h + 0h30 Horaire total : 2 heures |
Coût : 0 euros | Sécurité : RAS |
| Outils numériques et ressources | |
|---|---|
| ordinateur ou tablette ou smartphone | Image pc tablette smartphone
|
| logiciel Mesurim2 et la banque d’images |

image Mesurim2
|
| QR code | 
qr code
|
| Pearltrees | 
pearltrees
|
| Handi-accessibilité |
|---|
| descriptif audio |
Déroulement détaillé
Les élèves arrivent en classe de terminale avec des notions sur la méiose et des connaissances de génétique. Pour aborder les lois de Mendel, les élèves ont travaillé sur des modèles 3D d’épis de maïs disponibles dans la banque d’image de Mesurim2.

Parmi les divers organismes modèles disponibles, le maïs possède plusieurs caractéristiques pour illustrer les principes génétiques. Celles-ci incluent :
- une large gamme de mutants clairement visibles pour divers caractères, en particulier les caractères du grain.
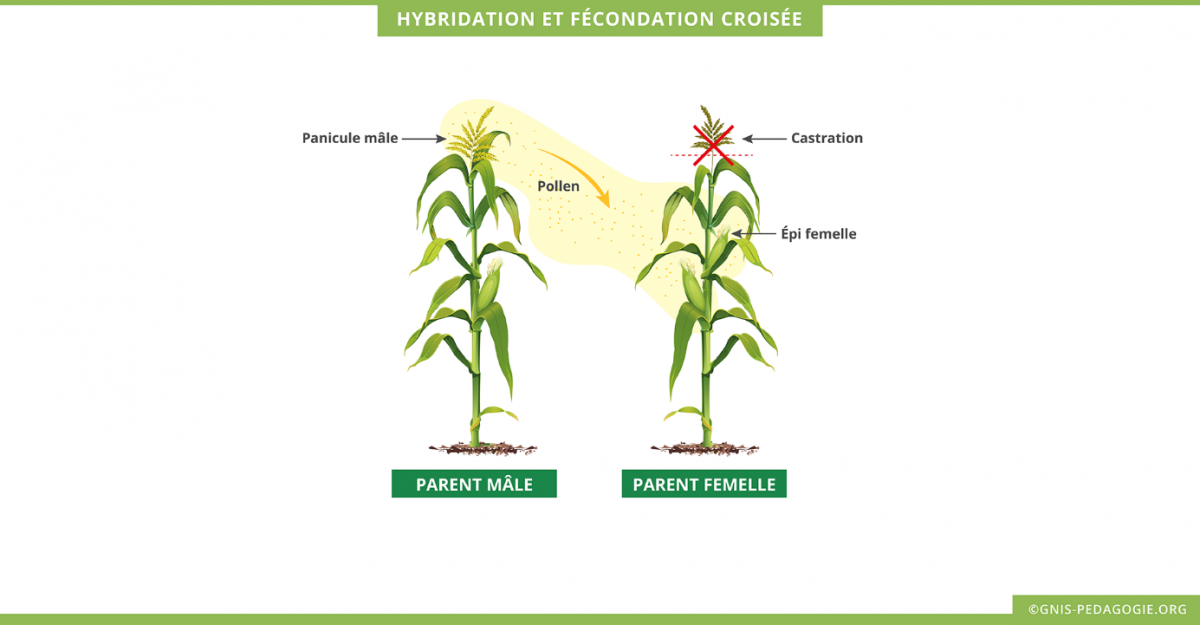
- les fleurs mâles et femelles, qui sont portées sur des structures séparées et facilitent les croisements. Le maïs est une plante monoïque.
- les grains sur un même épi, qui montrent une ségrégation génétique et représentent une population d’individus. La fleur de sexe mâle chez le maïs est la « panicule » à l’extrémité haute de la plante, sa fleur de sexe femelle est le futur « épi ».
- la séparation des graines disposées en rangées sur un épi, qui peut être utilisée pour illustrer des concepts statistiques tels que les événements aléatoires, l’échantillonnage et la probabilité.
Activité 1
La notion de croisement lois de Mendel
– Première loi : Loi d’uniformité des hybrides de première génération
Comme les gamètes sont issus de la méiose, ils ne possèdent qu’un seul allèle correspondant à l’un des deux chromosomes de l’individu "parent". Pour les lignées pures, c’est-à-dire les individus homozygotes récessifs ou dominants, les gamètes sont tous identiques puisqu’ils possèdent le même allèle.
Si l’on croise deux individus d’une même espèce homozygotes l’un dominant l’autre récessif, tous les descendants de la première génération, qui sont appelés hybrides F1, sont identiques relativement à ce caractère, c’est-à-dire tous hétérozygotes.
– Deuxième loi : loi de ségrégation des caractères dans la génération F2 ».
Pour un individu F1 hétérozygote, les deux allèles pour un même gène se séparent lors de la formation des gamètes (méiose) de sorte que 50% des gamètes obtiennent l’un des allèles et 50% des gamètes ont l’autre allèle.
À partir de deux individus F1, on obtient une génération F2 dans laquelle on trouve les deux versions du caractère dans des proportions bien définies : trois descendants au même phénotype (1 RR homozygote + 2 Rr hétérozygotes) et un descendant d’un autre phénotype (rr homozygote).
| À partir de l’observation des résultats de croisements chez le maïs, sur le caractère de la couleur des grains (noir ou jaune) et en vous aidant des connaissances actuelles en génétique, valider et justifier les résultats prévus par les lois de Mendel. |
| Précisions Les observations des phénotypes des grains de maïs correspondent dans ce cas à des observations d’une population diploïde, mais il ne s’agit pas d’une population d’embryons. Il est possible d’adapter les informations et la précision des caractéristiques du maïs en fonction du niveau de la classe. Il existe une double fécondation chez les plantes à fleur, une qui va donner le futur embryon et l’autre qui va donner l’albumen, tissu nourricier de l’embryon. Timelapse de 7 heures montrant le guidage des tubes polliniques (gris) Attraction tube pollinique
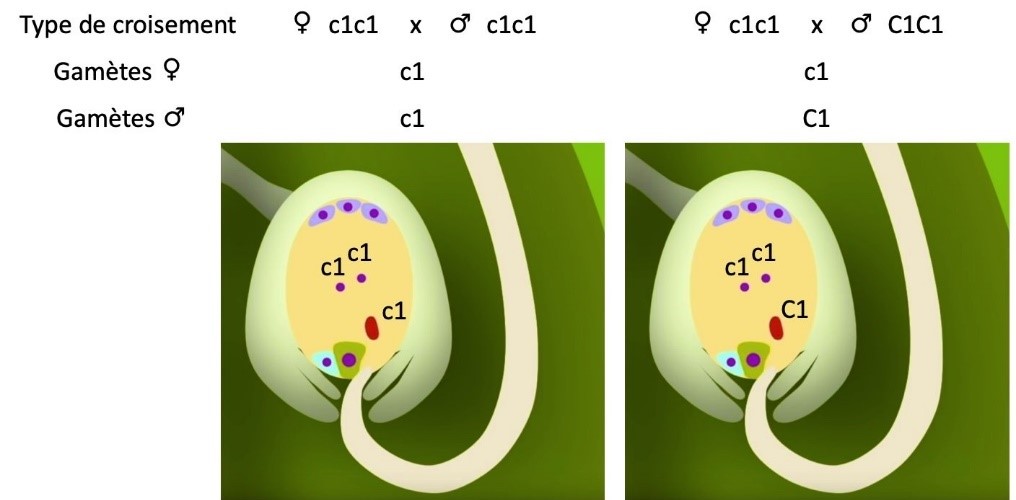
Schéma double fécondation
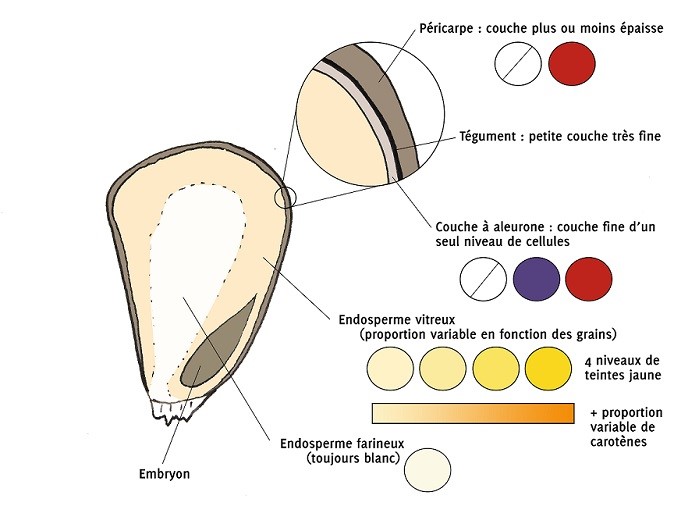
Schéma d’une coupe de grain de maïs avec les structures à l’origine de la couleur de l’albumen
|
Les élèves ont à disposition un lien vers le modèle directement disponible via Pearltrees et QR code si le modèle est présent dans la banque, Il y a également la possibilité d’ajouter des modèles 3D au format .glb dans Mesurim2. Les liens peuvent être modifiés afin de cacher les informations et les descriptifs de modèles.

| Ils ont également un lien vers le tutoriel de l’outil de comptage réalisé par Philippe Cosentino. |
Ils ont accès aux parents de lignées pures de couleur noire (R1//R1) et de couleur jaune (r1//r1) et au phénotype des individus F1.
| Dans le détail : Il s’agit d’un monohybridisme sur un caractère de la couleur noir/jaune avec :
|
Les comptages sont réalisés sur 3 lignes maximum et sont ensuite mutualisés via un tableur.

| Résultat total du croisement P1 et P2 | noir 100% | jaune 0% |
| Résultats total autofécondation F1 | noir 260 (76%) | jaune 82 (24%) |
Réalisation de l’échiquier de croisement et validation des lois de Mendel.
Modèle d’autofécondation avec 2 gènes pour la 3ème loi de Mendel, à venir
Activité 2
À la suite du TP sur les brassages intra et interchromosomique avec les drosophiles, cette activité a été proposée en classe entière et à distance afin d’approfondir la notion.
| Nous avons déjà vu les principes de Mendel avec l’étude de la couleur des grains de maïs et les brassages génétiques sur les mouches en cours et en TP. Nous allons essayer ranger les épis qui trainent dans le laboratoire, depuis des dizaines d’années. La seul indication qui reste est l’étiquetage des boites « parents », « F1 » et « F2 ». Plusieurs étiquettes avec des caractéristiques génétiques ont été retrouvées mélangées dans les boites. Certains épis sont issus d’un monohybridisme, d’autre d’un dihybridisme (les gènes de couleur et de forme peuvent être liés ou non). 
étiquette R//R x r//r

étiquette Y//y x y//y

étiquette ShC//ShC x shc//shc

Etiquette Su//Su R//R x su//su r//r
|
| Précision : Selon le niveau des élèves : L’effet de xénie est défini comme l’effet direct d’un type de pollen (et donc du génotype paternel) sur le développement du grain. Par la double fécondation, 1/2 du génome de l’embryon et 1/3 du génome de l’albumen proviennent directement du pollen. Un pollen qui porte l’allèle dominant (et donc qui n’est pas issu de la même plante/lignée) va influencer les caractères du grain, on parle alors d’effet de xénie. Cet effet a été particulièrement étudié chez le maïs. En effet il peut être observé facilement sur un épi sur des caractères macroscopiques. On peut citer des caractères comme la couleur, la taille ou encore la composition du grain. Le maïs doux a été sélectionné pour ses grains au taux de sucre particulièrement élevé. Lors de la maturation le maïs non doux a un l’albumen rempli d’amidon (composé de parts variables d’amylose et d’amylopectine). Ces 2 molécules sont des polymères plus ou moins ramifiés de glucose. Cet assemblage est contrôlé par plusieurs processus eux-mêmes contrôlés par des gènes et des facteurs de transcription. Des mutations dans ces gènes qui contrôlent la production d’amidon modifient la teneur en amidon et en glucose de l’albumen. Ici on a un exemple avec les mutations qui provoquent des pertes de fonctions des gènes Sh2 (shrunken 2) et Su1 (sugary1 type). Métabolisme de production de l’amylose et de l’amylopectine dans l’albumen du grain de maïs 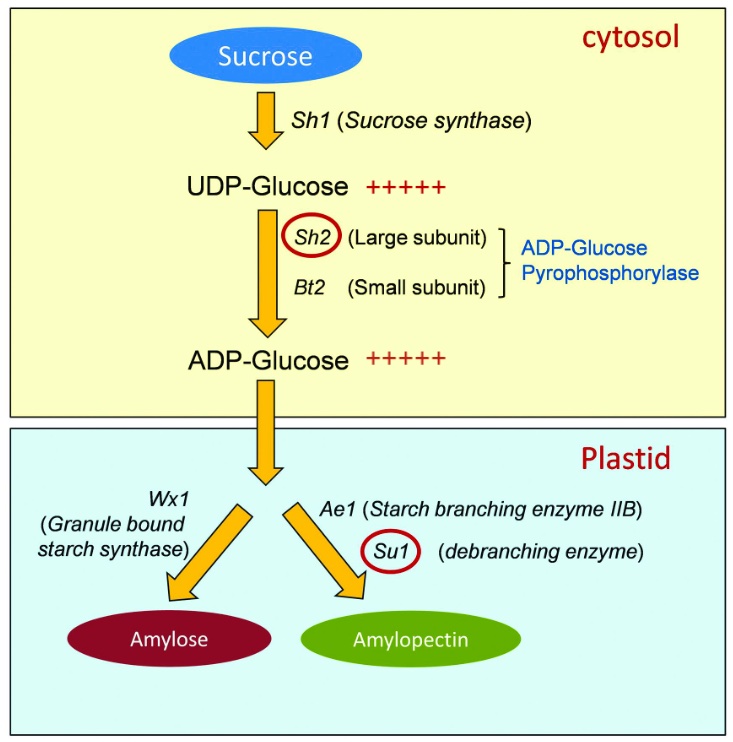
Schéma métabolisme et effet mutation sh et su
|
| Aide au comptage : Un tableau collaboratif de vos comptages est disponible pour augmenter la fiabilité des résultats. Pour chaque individu ne compter que 3 lignes de grains. |
Aide « lois » de Mendel :
|
| Aide croisement Rétrocroisement ou back cross Le premier croisement ou croisement test entre deux parents de lignées pures P1 x P2 donne la génération F1 qui permet de déterminer le caractère dominant d’un allèle, ici pour les gènes déterminant la couleur ou la forme du grain. Le deuxième croisement entre F1 et le parent double récessif, appelé génération F2, montre des phénotypes qui correspondent aux caractéristiques des génotypes des gamètes de F1. Dans le cas de gènes indépendants les résultats seront de ¼, s’ils sont liés, les résultats seront différents entre les combinaisons de parentaux et des combinaisons recombinés. Hybride = organisme issu du croisement de deux lignées « pures » différentes. Pour un caractère (monohybydisme), on croise des individus homozygotes pour l’allèle responsable de ce caractère. Lors de la méiose les cellules produites reçoivent un seul des deux allèles de chaque paire avec une probabilité équivalente. Pour deux paires d’allèles, quatre combinaisons d’allèles sont possibles pour les gamètes produits : Le brassage interchromosomique est observé avec plusieurs gènes portés par des chromosomes différents. Les 4 combinaisons d’allèles sont équiprobables : 25%. Les phénotypes parentaux sont plus probables que les phénotypes recombinés par brassage interchromosomique et intrachromosomique, si les gènes sont liés. |
| Aide génétique des Maïs Tableau des gènes impliqués dans la forme et la couleur |
| gène | numéro de chromosome | Allèles | Rôle |
| SH1 | chromosome 9 | Sh1 Rond sh1 Plissé | synthèse du glucose endosperme |
| C1 | chromosome 9 | C1 violet rouge c1 sans couleur | endosperme transcription des anthocyanes |
| Y | chromosome 6 | Y1 jaune y1 blanc | endosperme synthèse de carotène |
| R1 | chromosome 10 | R1 violet rouge r1 sans couleur | endosperme synthèse d’anthocyane facteur de transcription |
| Su 1 | chromosome 4 | Su1 rond su1 ridé | endosperme production de sucre |
Les élèves ont accès comme la première activité à des liens directs dont toutes les informations des épis sont absentes.
Exploitation de modèles 3D d’objets réels : épis de maïs issus de croisements différents de parents homozygotes pour des caractères de couleurs et de formes
Travail sur navigateur internet via tablettes ou PC



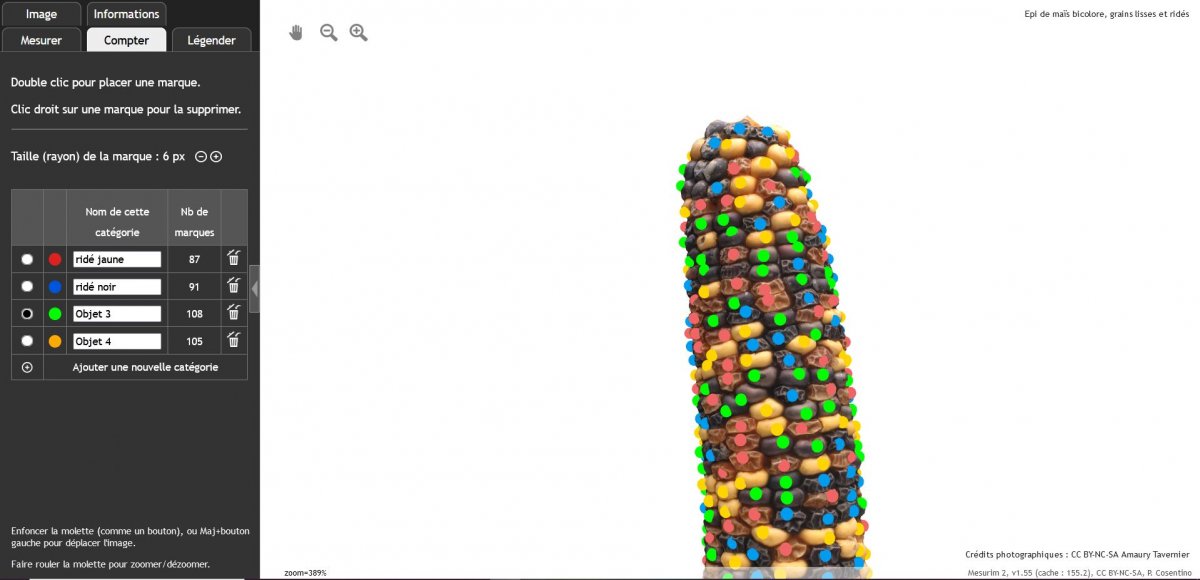
Résultats des croisements F2
| F2 2 gènes indépendants noir//jaune lisse//ridé | |
| Noir ridé | 98 (23.6%) |
| Noir lisse | 118 (28.5%) |
| Jaune ridé | 92 (22.1%) |
| Jaune lisse | 107 (25.8%) |
| F2 2 gènes liés noir lisse//jaune plissé | |
| Noir lisse | 107 (43.5%) |
| Noir plissé | 13 (5.3) |
| Jaune lisse | 11 (4.5%) |
| Jaune plissé | 115 (46.7) |
| Autofécondation noir//jaune | |
| Noir | 260 (76%) |
| Jaune | 82 (24%) |
| Autofécondation f1 2 gènes liés noir lisse//jaune plissé | |
| Noir lisse | 265 (61%) |
| Noir plissé | 50 (11.6%) |
| Jaune lisse | 9 (2%) |
| Jaune plissé | 105 (24.4%) |
Focus sur un outil : Mesurim 2
Mesurim2 est une application créée par Philippe Cosentino (académie de Nice), qui permet de réaliser des mesures de longueurs, d’angles, de surfaces, comptages, comparaisons ou d’observation en microscopies virtuelles sur des photographies, ainsi que des observations de modèles en 3D. Une banque d’images fournit des modèles partagés de façon collaborative par des collègues de SVT. Il est également possible d’intégrer ses propres photos ou celles des élèves.
| Lien direct vers le logiciel Mesurim2 |
|
image Mesurim2
|
| Lien vers le tutoriel de comptage réalisé par P. Cosentino - académie de Nice : |
Tutoriel comptage
|
| Lien vers Les caractéristiques crâniennes du genre Homo : crâniométrie virtuelle sur Mesurim2 (Philippe Cosentino) |
Les caractéristiques crâniennes du genre Homo : crâniométrie virtuelle sur Mesurim2
|
Avantages / Plus-values :
- Relier réel et virtuel
- Très nombreux outils pour les mesures avec des tutoriels intégrés
- Facilité d’utilisation
- Outil en ligne/gratuit
- Évolution constante
- Nombreux modèles déjà disponibles et libres
- Modèles rigoureux
Points de vigilance :
- Ne remplace pas l’étude du réel
Retours, analyse et pistes d’amélioration
| RETOUR DES IMPRESSIONS DES ÉLÈVES |
|---|
Retour direct : Facilité d’usage et d’exploitation de l’outil ainsi que de la prise en mains, malgré quelques bugs techniques. Usage de tous les supports, mais plus compliqué sur smartphone.
Situation de la classe
| classe de terminale de 34 élèves spécialité SVT et PC |
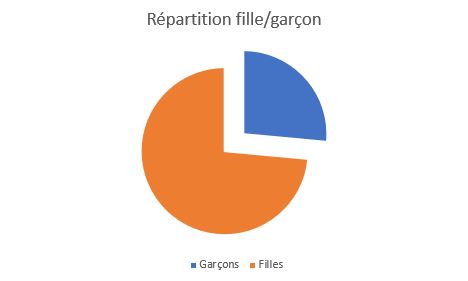
Répartition fille/garçon
|
La classe est très déséquilibré, avec une très large majorité de fille. Dans notre lycée, 60 élèves ont gardé la spécialité SVT sur 140 en première, avec environ 75% de fille. |
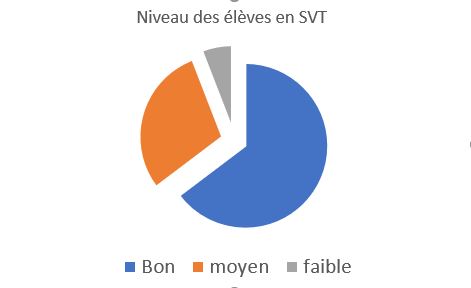
Niveau global en SVT
|
Le niveau global de la classe est très bon, avec une grande autonomie dans le travail et un groupe sérieux qui travail en collaboration. Quelques élèves sont en relative difficulté depuis la 1ère. |
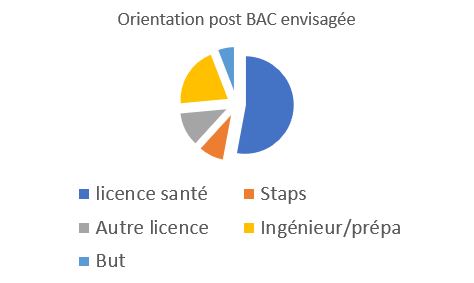
orientation post bac
|
La classe est très orienté vers les métiers du médical et plus particulièrement Pass et Las. Le choix de faire une classe exclusivement en spécialité SVT et PC polarise encore plus ces choix |
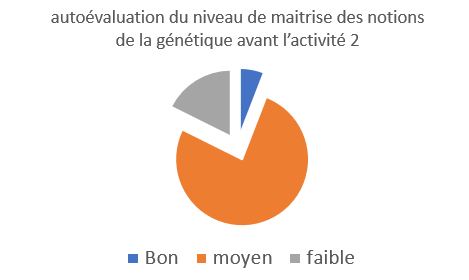
autoévaluation du niveau de maitrise des notions de la génétique avant l’activité 2
|
Ici, le niveau est déclaratif, suite au TP avec les drosophiles, les élèves semblent ne pas avoir confiance dans ce qu’ils ont déjà fait. Ce sentiment est global sur la classe, qui a tendance à dévaloriser ses capacités. |
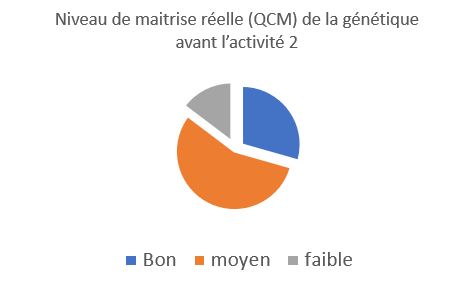
Niveau de maitrise réelle (QCM) de la génétique avant l’activité 2
|
Les élèves ont réalisé un QCM sur les définitions et notions abordées lors des cours et TP précédents. Les résultats montrent une certaine maitrise des compétences et connaissances, surtout cette maitrise est bien supérieur à l’autoévaluation. |
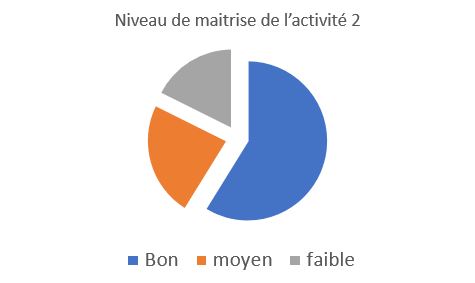
Niveau de maitrise de l’activité 2
|
Le résultats présentent ici le niveau du compte-rendu de l’activité 2, les attendus correspondaient à la mise en relation correct des étiquettes avec la présentation des résultats sous forme de tableaux, ainsi que les échiquiers de croisement correspondants et une conclusion. Les élèves semblent avoir bien mis en relation les notions et avoir utilisé les bons termes. Un groupe d’élèves n’a pas pu ou n’a pas réussi à mettre en relation les résultats et a réalisé des échiquiers erronés |
| ANALYSE ET ÉVALUATION DU DISPOSITIF | |
|---|---|
| Plus-values dégagées |
|
| Difficultés rencontrées |
|
| Pistes d’amélioration |
|
Source : https://www.semae-pedagogie.org/GNIS-pedagogie.org, https://www.isgpb.org/article/genet..., Yann Duroc, Nick Desnoyer

