
La métaphysique est la recherche du pourquoi ultime, ce qu’Aristote nommait la cause première. Elle se distingue ainsi de la science qui est plutôt une recherche du comment (bien que des scientifiques n’acceptent pas cette limite). Elle fait ainsi écho à des interrogations fondamentales de l’homme poussé dans la quête d’un sens, interrogations profondes auxquelles la science ne peut répondre car cela dépasse son champ d’application (Nous en reparlerons en évoquant le positivisme d’Auguste Comte et le scientisme).
Un philosophe (étymologiquement au moins) aime la sagesse. Le discours philosophique peut se nourrir des résultats de la science, et il peut à son tour interroger la science et la pousser vers de nouvelles voies. L’époque des Lumières où les champs du savoir étaient moins cloisonnés qu’aujourd’hui est riche de tels exemples. Les travaux de Goethe sur l’archétype foliaire ou ceux de Geoffroy Saint Hilaire sur l’unicité de plan d’organisation sont ainsi inspirés par la Naturphilosophie allemande.
Toutefois si la philosophie peut proposer de nouvelles idées (de nouvelles hypothèses pourrait-on dire), ce sont les seuls faits, qui doivent arbitrer la pertinence de ces idées pour le scientifique. Les idées doivent céder devant les faits, faute de quoi on retrouve l’esprit de systèmes qui prévalait avant les Lumières, lorsque l’étude de la nature se faisait sur un mode spéculatif qui se préoccupait peu de se confronter aux données du réel. La science ne peut trouver ses explications que dans le cadre de la nature.
Mais il n’est pas si évident d’être complètement objectif lorsqu’on analyse des résultats aussi concrets soient-ils. Cette objectivité relève en grande partie d’une mythologie de la science, d’un idéal que notre nature humaine, notre cerveau pas parfaitement logique car fruit de l’évolution, ne nous permettent pas d’atteindre. Il peut toujours exister une influence extérieure aux données, liée à notre culture, à notre conception du monde qui ne se construit pas que de science. La pensée du scientifique ne peut pas se couper du reste de son esprit. Il ne faut jamais négliger que la science n’avance pas indépendamment sous la seule influence des faits et des progrès techniques. Parce qu’elle est œuvre humaine, elle subit nécessairement une influence du cadre social, des modes de pensée contemporains, de la philosophie dominante de l’époque. Ce n’est sans doute pas un hasard si la notion d’évolution du monde vivant est née à l’époque des Lumières, dans un monde intellectuel et dans des sociétés en plein changement. De même pour la théorie darwinienne née dans l’Angleterre industrielle et libérale : on connait l’influence qu’a eu la lecture de Malthus sur Darwin. On pourrait aussi souligner l’influence d’Adam Smith.
C’est pourquoi il est pertinent même pour qui se veut d’abord scientifique de se pencher sur l’histoire des idées et de les suivre dans une histoire des sciences car cela permet de reconnaître ce qui a pu orienter ou déformer l’interprétation des faits.
Il m’est apparu aussi simplement important d’essayer de définir ma propre position philosophique pour répondre à l’injonction socratique « Connais-toi toi-même ».

Platon, Aristote (détail de la fresque de Raphaël L’École d’Athènes, Salle des signatures, Vatican, Rome)... et Charles Darwin
I. La théorie de l’évolution en résumé et l’évolution de la théorie
II. Hasard et finalisme
III. Vitalisme et mécanisme
IV. La place de la Science, Morale et Religion
V. Enseigner l’évolution de la lignée humaine
I. La théorie de l’évolution en résumé et l’évolution de la théorie
Il faut tout d’abord bien séparer l’évolution de ses mécanismes. L’évolution est un fait avéré, observé et bien documenté, par les fossiles, par l’anatomie comparée, par les études moléculaires. Invalider Darwin ne reviendrait de toute façon pas à invalider l’évolution.
Les mécanismes eux font l’objet d’une théorie qui s’affine depuis plus de 150 ans, depuis Darwin, se complexifie et se complète, résistant d’une façon remarquable aux nouvelles observations et remises en cause, tout cela étant preuve de sa vitalité.
Avant de discuter des pendants philosophiques, commençons par rappeler en termes simples ce que nous dit la théorie de l’évolution, qui découle des idées de Darwin, enrichies ensuite par les connaissances notamment en génétique, afin de bien poser ce qui sera discuté plus tard.
Il ne s’agit pas d’un cours exhaustif sur l’évolution. Chaque partie nécessiterait de plus amples explications.
Pour en savoir plus sur les principaux thèmes :
– un dossier sur le site Vie
– un dossier sur le site du CNRS
Pierre Henri Gouyon résume ainsi l’évolution :
« Les informations contenues dans l’ADN sont reproduites de génération en génération ; ce faisant, elles varient sous l’effet des mutations et des recombinaisons et sont triées par la sélection naturelle. Ce processus, répété sur quelques milliards d’années, a donné naissance aux formes vivantes actuelles. A chaque pas la mutation se fait au hasard (c’est-à-dire qu’elle n’est pas dirigée vers un résultat, la direction n’est donnée qu’a posteriori par la sélection. » [1]
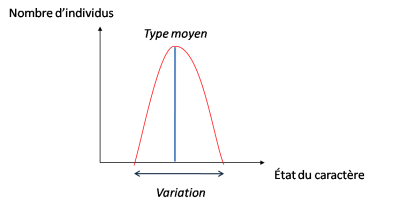
La théorie de l’évolution peut donc se résumer par le couple variation/sélection.
– des variations apparaissent au hasard dans la population
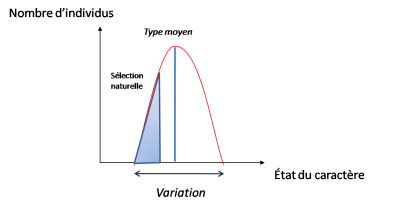
– dans un milieu donné certaines se révèlent favorables. Elles permettent une meilleure survie, et donc une meilleure reproduction, des individus qui les possèdent. Les autres sont éliminés ou ont un accès plus difficile à la reproduction.
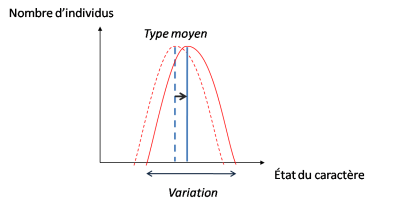
Le type moyen est ainsi déplacé.
Ce tri parmi les variants correspond à la sélection naturelle de la théorie de Darwin telle qu’on la trouve dans L’origine des espèces.
Pour Darwin, la variation est isotrope, c’est-à-dire qu’elle n’est nullement orientée et se fait dans toutes les directions possibles. La sélection naturelle intervient ensuite et est l’agent essentiel qui va faire évoluer les espèces. Son niveau d’action est l’organisme et elle sélectionne des caractères, c’est-à-dire un phénotype, ensemble des caractères de l’individu.
Plus tard, lorsque la théorie darwinienne a été couplée avec la redécouverte de Mendel augmentée du concept de mutation introduit par Hugo De Vries (XIXe siècle), la variation, dont l’origine est inconnue pour Darwin, prend corps au niveau de l’ADN, avec les modifications qui peuvent toucher cette information génétique et qui seront de mieux en mieux connues et comprises tout au long du XXème siècle, et qui sont encore des sujets d’étude féconds et importants aujourd’hui. La génétique des populations va aussi être un apport important en suivant précisément les changements qui se produisent dans les fréquences alléliques au fil du temps. Ils constituent la microévolution, distinguée de la macroévolution, menant à l’apparition de nouvelles espèces ou de nouveaux taxons au-dessus de l’espèce.
Une vision plus complète des mécanismes de l’évolution nécessite de donner quelques précisions, tant au niveau des mécanismes de la variation qu’au niveau de la sélection.
- La variation au niveau génétique :
- Les innovations génétiques
- Les mutations
- Les innovations génétiques
Lorsque l’on pense variation, on pense souvent d’abord à la mutation génétique ponctuelle : il se produit un changement dans la séquence de nucléotides de l’ADN. Ces changements surviennent au moment de la réplication de l’ADN et sont finalement de simples erreurs de copie, qui ont échappé aux systèmes de vérification et de correction qui existent pourtant.
Ces accidents peuvent être de différentes sortes : remplacement, ajout ou délétion d’un ou plusieurs nucléotides.
Les conséquences en sont diverses, fonction des propriétés du code génétique (redondance qui protège contre les effets de ces mutations, existence de codons non sens) et du mécanisme de la traduction.
Il se crée ainsi de nouveaux allèles, c’est-à-dire de nouvelles versions des gènes, qui correspondront à autant de versions légèrement différentes du même caractère. Ce sont ces versions différentes du caractère qui seront soumises à la sélection naturelle.
Pour en savoir plus : Mutation et nouveauté par Jacqueline Laurent, bibliothèque Pour La Science : L’évolution
Mais il peut arriver d’autres « accidents » à l’ADN.
-
-
- Transposition
-
Une partie de l’ADN possède la capacité de se transposer. Appelés éléments transposables, ces fragments d’ADN ont la capacité de se multiplier et de se déplacer dans le génome. Des copies vont ainsi s’insérer ailleurs, au hasard.
Ceci peut s’interpréter comme étant un vestige de l’origine virale de notre génome. Cette transposition reproduirait une partie du cycle d’un virus qui autrefois aurait infecté la cellule et qui pour une raison ou une autre n’en serait plus ressorti, transmis de génération en génération avec le génome, en faisant partie.
En s’insérant ainsi au hasard, il peut arriver qu’une copie se place au sein d’un gène et donc l’inactive.
Il arrive aussi que des gènes soient emportés et que l’on se retrouve ainsi avec plusieurs copies du même gène. Dans un premier temps ces copies codent pour la même information mais les mutations ponctuelles intervenant, leurs séquences vont diverger et il pourra arriver que l’une de ces séquences soit retenue par la sélection car apportant une nouvelle fonction. Se constituent ainsi des familles multigéniques, familles de gènes dont les rôles ne sont pas identiques mais qui sont issus d’un même gène ancestral. Les pigments rétiniens ou les diverses hémoglobines en sont des exemples classiques.
Pour en savoir plus : Séquences d’ADN mobiles et évolution du génome par Claude Bazin, Pierre Capy, Dominique Higuet et Thierry Langin, bibliothèque Pour La Science : L’évolution
-
-
- Les réarrangements chromosomiques
-
A une échelle supérieure, il peut arriver que lors de la formation des gamètes, c’est-à-dire les cellules reproductrices (classiquement ovule et spermatozoïde) des chromosomes se scindent, fusionnent, échangent des morceaux, ou même « oublient » de se séparer.
Chez de nombreuses espèces, les chromosomes sont présents en un nombre pair d’exemplaires, souvent deux. Ces chromosomes qui vont par deux la plupart du temps sont dits homologues. Ils portent les mêmes gènes mais pas nécessairement les mêmes allèles de ces gènes (l’individu est homozygote lorsqu’il possède deux fois le même allèle, hétérozygote s’il possède deux allèles différents pour un même gène). L’un vient de l’organisme maternel, l’autre de l’organisme paternel.
Pour former les gamètes, il y a séparation de ces paires de chromosomes homologues lors de la méiose (elles seront reconstituées lors de la fécondation).
Au cours de cette méiose, il y a une étape nécessaire d’appariement des chromosomes homologues, au cours duquel ils vont s’attacher en plusieurs points et vont pouvoir échanger des morceaux (on parle de recombinaison et de brassage intra chromosomique). Ce sont des « accidents » dans ces mécanismes qui vont entraîner les réarrangements évoqués plus haut.
L’accident le plus extrême est lorsqu’il n’y a pas séparation des chromosomes homologues. Lorsque cela touche une seule paire de chromosomes, on parle de trisomie. Il arrive aussi que toutes les paires restent ensembles. On a alors un organisme dit polyploïde qui va posséder chaque chromosome non pas en deux mais en 4 exemplaires, ou six, huit…, (un nombre nécessairement pair pour qu’il puisse y avoir appariement lors de la méiose).
L’hybridation, croisement entre espèces, peut aussi produire des polyploïdes.
Ces réarrangements sont des macromutations, par opposition aux micromutations que sont les mutations ponctuelles.
Pour en savoir plus :
– Une autre manière de créer des espèces par Bernard Dutrillaux dans le dossier La Recherche mai 2007 L’évolution
– Chromosomes, systématique et évolution par Vitaly Volobouev, bibliothèque Pour La Science : L’évolution
– Une conférence de l’UTLS : Végétaux, diversité, hybridation de Jacques Mugnier
-
- Les transferts horizontaux
Des gènes peuvent se transmettre directement d’un organisme à un autre, même d’espèces différentes. Cela constitue un transfert horizontal, par contraste avec les transferts verticaux qui correspondent à la transmission héréditaire. Ces transferts horizontaux existent en particulier dans le monde microbien, avec le phénomène de conjugaison et la transformation bactérienne, et viennent bouiller les arbres phylogénétiques que l’on peut y établir. Le hasard des rencontres intervient donc ici aussi.
Pour en savoir plus : Les arbres du vivant : source de discorde - Chloé Terras dossier Pour La Science Où est née la vie ?
-
- Transmission à la descendance
Toutes ces variations au niveau génétique vont potentiellement créer de la nouveauté au niveau des caractères. Que faut-il pour que cela soit transmis à la descendance ?
La transmission est directe lors d’une reproduction asexuée comme la division d’une bactérie.
Chez les pluricellulaires intervient la séparation du soma et du germen, formulée par August Weismann.
Toutes ces modifications du génome pour entrer dans le jeu de l’évolution, pour être soumises à une sélection, doivent être transmissibles à la descendance. Il faut donc qu’elles affectent des cellules de la lignée germinale, cellules à l’origine des gamètes et donc susceptibles de former la génération suivante. Une mutation affectant une cellule somatique, non à l’origine de gamètes (par exemple les cellules des poumons, du foie, du cœur…), est condamnée à disparaître avec l’organisme.
Mais pour être transmise à la génération suivante, il ne suffit pas que la modification génétique touche une cellule germinale, il faut en plus que celle-ci soit à l’origine du gamète impliqué dans la fécondation.
- La sélection
-
- La sélection naturelle
La sélection naturelle a été maintes fois observée et est un fait avéré. On en trouvera de nombreux exemples, tels que l’aspect des Guppies qui varie en fonction de la présence des prédateurs. Elle a été observée dans la nature, comme sur les pinsons de Darwin, sans parler du fameux et controversé exemple de la phalène du Bouleau.
Pour en savoir plus :
– Le papillon de la discorde - Michael E.N. Majerus ; Le bec du Pinson des Galapagos - Elisabeth Pennisi, dossier La Recherche mai 2007 L’évolution
– La sélection naturelle et les pinsons de Darwin - Peter Grant, bibliothèque Pour La Science : L’évolution
Les expériences de Teissier et l’Héritier avec leurs cages à popultions de Drosophiles ont permis de montrer cette réalité de façon expérimentale.
Pour en savoir plus : Evolution et génétique des population - Jean Génermont, bibliothèque Pour La Science : L’évolution
-
- La sélection sexuelle
La sélection sexuelle a été définie par Darwin pour expliquer certains traits qui semblent plus entraver qu’aider la survie de l’individu, telle que la queue du Paon.
Il s’agit ici non d’une survie différentielle, mais d’un accès différentiel à la reproduction. Il s’agit d’une compétition entre les individus pour l’accès aux représentants de l’autre sexe, par la séduction à l’aide certains attributs, comme la queue du Paon, ou par des combats entre prétendants.
- La spéciation.
La spéciation correspond à l’apparition d’une nouvelle espèce. On définit le plus couramment en biologie une espèce par une communauté reproductive. Deux individus appartiennent à la même espèce s’ils peuvent avoir une descendance fertile. Cette définition est imparfaite, en particulier chez les Végétaux, parce qu’elle est artificielle. De nombreuses espèces, bien distinctes sur la base de leurs caractères, peuvent donner des hybrides fertiles, bien que cette fertilité soit souvent imparfaite.
Pour en savoir plus : Qu’est-ce qu’une espèce ? - Jean Génermont, bibliothèque Pour La Science : L’évolution
Dans les faits, la spéciation est donc établie lorsqu’il n’y aura plus d’échange génétique possible entre deux communautés d’individus, pour des raisons chromosomiques, morphologiques ou de comportement.
Beaucoup de cas d’hybridation avec une descendance fertile peuvent s’interpréter comme une spéciation imparfaite, pas encore terminée. Plus la spéciation sera avancée, plus la fertilité des hybrides sera réduite. Puis il n’y aura plus de développement embryonnaire possible, plus de fécondation, et enfin plus d’accouplement. Entretemps, avant de parler de deux espèces différentes, on parlera de variétés ou de sous espèces, grande difficulté depuis des siècles pour les taxinomistes qui établissent les classifications et doivent décider de leur statut.
Si on reprend notre couple variation/sélection, on voit au sein d’une population les fréquences des caractères changer au fil du temps. Le type moyen de l’espèce à un temps t1 ne ressemblera plus au type moyen de l’espèce au temps t2. On a alors ce que l’on appelle une anagenèse : une espèce se transforme en une autre espèce. C’est ainsi que Lamarck voyait l’évolution, bien qu’il ait dû modérer, suite à ses observations, cette vision linéaire.
Mais ce n’est pas tout à fait ce que les archives fossiles nous révèlent. Les fossiles montrent une multiplication des lignées, ce que l’on appelle une évolution buissonnante. Une espèce va donner plusieurs espèces par une divergence des lignées. C’est une des nouveautés introduites par Darwin par rapport à Lamarck. On parlera de cladogenèse.
On distingue deux modes de spéciation
-
- Spéciation allopatrique
La population se trouve scindée en deux par un obstacle naturel (montagne, océan…). Des deux côtés de l’obstacle les conditions ne sont pas les mêmes, donc la sélection naturelle ne retiendra pas les mêmes caractères. Chaque population évoluera différemment de son côté et finira par constituer une nouvelle espèce.
Cela peut s’observer aussi sans même qu’il y ait d’obstacles en bordure de l’aire de répartition de l’espèce, c’est-à-dire en bordure du milieu auquel les membres de l’espèce sont adaptés. Ceux qui vivent en périphérie sont au contact d’autres milieux, à des conditions plus changeantes. Dans les faits, c’est en périphérie des aires de répartition que l’on peut observer le plus grand nombre de variétés. On l’observera par exemple au niveau des littoraux, dont la végétation est très caractéristique mais aussi souvent cousine de la végétation plus éloignée du bord de mer.
Sans même parler d’environnements différents et de sélection différente, la dérive génétique seule pourra faire diverger les deux espèces, puisque la grande loterie de la reproduction ne donnera pas le même résultat des deux côtés de l’obstacle.
Cela sera d’autant plus vrai que l’une des populations sera petite. La dérive sera d’autant plus forte, puisque le nombre de tirages sera d’autant plus faible. A cela s’ajoutera le fait que cette population réduite n’aura emporté avec elle qu’une partie de la diversité génétique de la population initiale. Les tirages possibles ne sont donc pas les mêmes.
-
- Spéciation sympatrique.
Dans ce cas les deux espèces divergent à partir de l’espèce ancestrale sur un même territoire. Les cas qui sont connus correspondent le plus souvent à des réarrangements chromosomiques. Pour que le réarrangement soit fixé, il faudra qu’il n’empêche pas la reproduction de l’individu, ne perturbe pas trop la méiose. Il pourra exister ainsi à l’état hétérozygote jusqu’à ce que les hasards de la reproduction fassent naitre un homozygote avec deux exemplaires remaniés du chromosome touché.
La transmission est plus simple chez les espèces pratiquant l’autofécondation. C’est pourquoi ce mode de spéciation est plus courant chez les végétaux, chez lesquels on connaît de nombreux polyploïdes.
La spéciation allopatrique était autrefois considérée comme le mode majeur de spéciation. Mais il semble que la spéciation sympatrique soit plus courante qu’on ne pensait.
Pour en savoir plus : Les preuves de la spéciation sans frontières - Pierre Capy, dossier La Recherche mai 2007 L’évolution
Il reste de nombreuses questions et pistes ouvertes dans la théorie de l’évolution, ce qui n’est pas le signe de sa sénescence mais celui de sa vitalité.
- Sur la sélection naturelle
Depuis Darwin, même alors que la sélection naturelle est reconnue, la question se pose de déterminer ce qu’est son pouvoir réel sur l’évolution : est-elle le moteur principal ou y en a-t-il d’autres qui la dépassent ?
La sélection naturelle n’a pas été complètement inventée par Darwin mais pour d’autres auteurs elle agissait non pas comme une force diversifiante, qui crée de la nouveauté, mais comme une force stabilisante, c’est-à-dire éliminant tous ceux qui sont trop éloignés du type moyen. Cette conception s’inscrit dans une pensée essentialiste où tous les membres d’une même espèce partagent la même essence, qui est invariable. Cette essence correspond au type moyen de l’espèce, un archétype et dans laquelle la variation est un accident, une monstruosité.

Pendant toute une partie du XXème siècle, les évolutionnistes ont montré un peu trop de zèle dans l’emploi de la sélection naturelle pour leurs explications et finissaient par brosser un tableau dans lequel tout était pour le mieux parce que tout dans les êtres vivants avait été sélectionné, façonné par la sélection naturelle.
C’est pourquoi ils se sont vus qualifiés de panglossiens, du nom de la caricature de Leibniz faite par Voltaire dans Candide. Ils finissaient par adopter un raisonnement proche de celui de Paley dans sa théologie naturelle, Dieu étant remplacé par la sélection naturelle. On parle de pansélectionnisme.
Or la sélection naturelle n’est pas toute puissante :
-
- La théorie neutraliste et la dérive génétique
Motoo Kimoora a proposé qu’un certain nombre d’allèles ne soient ni favorables ni défavorables pour l’organisme. Ils sont dits neutres et échappent donc à la sélection naturelle. Leur transmission d’une génération à l’autre se fera donc de façon aléatoire. Leur devenir dépendra de la proportion de ces allèles dans la population et des hasards de la reproduction. Il pourra se produire une dérive génétique, c’est-à-dire que les proportions de ces allèles neutres dans la population vont varier au cours du temps, selon les hasards de leur transmission.
Imaginons un dé à 6 faces numérotées. 2 faces avec le numéro 1, 4 faces avec le numéro 2. La probabilité de sortie de chaque numéro est de 1/6 multiplié par le nombre de faces portant ce numéro. Mais cette probabilité ne deviendra réalité que si on procède à un nombre suffisant de tirages. Si on ne fait que 6 tirages, le résultat sera différent. Disons que sortent trois fois le 1 et trois fois le 2.
Le dé pour la seconde génération portera trois fois le 1 et trois fois le 2. Les proportions se sont donc modifiées sous l’effet du hasard.
En pratique, la reproduction est comme un tirage au sort. La reproduction sexuée correspond à la rencontre de deux cellules reproductrices, qui appartiennent à deux individus. Il y a donc une grande part de hasard dans cette double rencontre : rencontre des individus parmi toute la population, rencontre des deux cellules reproductrices parmi toutes celles qui sont produites par les deux individus.
La présence des allèles dans la population varie donc en fonction du hasard lors de la transmission d’une génération à l’autre, si ces allèles correspondent à des caractères échappant à la sélection naturelle.
Cette dérive sera d’autant plus marquée que la population aura un faible effectif, c’est-à-dire que le nombre de tirage sera faible. On parlera d’effet fondateur.
Pour en savoir plus :
– La théorie neutraliste de l’évolution moléculaire par Motoo Kimura, bibliothèque Pour La Science : L’évolution
– Un article sur le site Vie : La dérive génétique
-
- La cosélection
Les gènes ne sont pas indépendants les uns des autres mais sont les uns à la suite des autres sur le fil de l’ADN. Lorsqu’un allèle est sélectionné, ses voisins proches le seront aussi même s’ils n’apportent pas d’avantage particulier à leur porteur. Il faut une recombinaison, c’est-à-dire un échange entre deux chromosomes homologues lors de la méiose, pour que deux allèles se retrouvent séparés. Or plus deux allèles sont proches, plus la probabilité qu’ils soient séparés est faible (c’est sur cette base que les équipe de Morgan ont établi les premières cartes génétiques au début du XXème siècle).
- Sur la variation :
Pour Darwin, la variation est abondante et isotrope : elle se fait dans toutes les directions possibles.
Un débat antérieur à Darwin oppose le structuralisme et le fonctionnalisme. Le premier donne la primauté à la structure, le second à la fonction. Pour le premier, l’organe crée la fonction alors que pour le second c’est la fonction qui crée l’organe. Le transformisme de Lamarck avec l’effet de l’usage et du non usage est de type fonctionnaliste.
Un épisode fameux de ce débat a été l’opposition entre Cuvier et Goeffroy Saint Hilaire, l’un étant fixiste et l’autre transformiste. La raison pour laquelle Cuvier ne pouvait accepter le transformisme de Lamarck et l’unicité de plan d’organisation de Goeffroy qui faisait primer la structure sur la fonction, c’est parce qu’il voyait dans l’anatomie un arrangement bien précis dans lequel chaque organe joue son rôle en relation avec les autres. Cette harmonie dans la construction de l’organisme, ce lien qu’il voyait entre toutes les parties, était à la base des raisonnements qu’il suivit pour reconstituer les organismes fossiles incomplets :
« En un mot, la forme de la dent entraîne celle du condyle, celle de l’omoplate celle des ongles, tout comme l’équation d’une courbe entraîne toutes ses propriétés. »
Pour Cuvier, fonctionnaliste, c’est la fonction qui prime et si on retrouve les mêmes arrangements entre plusieurs organismes, c’est à cause de contraintes fonctionnelles.
Il ne voyait donc pas comment l’un pouvait se transformer sans perturber de façon délétère tout l’équilibre délicat de l’organisme.
On peut traduire cette critique par le terme de contrainte : toutes les variations ne sont pas possibles dans un organisme. Il y a des contraintes liées au fonctionnement de l’organisme, ce à quoi on peut ajouter des contraintes liées à son développement.
Ainsi il est possible que la variation se fasse plutôt dans une direction privilégiée.
Une autre critique de la variation isotrope est la lecture de tendances lorsque l’on suit les caractères dans une lignée. (Cette lecture de tendance est elle-même souvent critiquable car elle est marquée par la subjectivité dans le choix des caractères). L’interprétation darwinienne classique lorsque l’on voit un caractère changer dans la même direction sur une longue durée est d’invoquer une sélection qui est restée la même.
D’autres veulent y voir les signes d’un déterminisme interne, d’une évolution guidée de l’intérieur. Les plus extrêmes, que l’on peut qualifier de finalistes, voient un but inscrit dans l’évolution.
Les tenants de ce déterminisme interne ont souvent utilisé l’étude du développement embryonnaire comme argument, rapprochant ontogénie et phylogénie en reprenant la théorie de la récapitulation qui postule que le développement embryonnaire répète l’évolution. En hiérarchisant les êtres vivants, l’évolution se fait par ajout d’étapes du développement. Pour eux l’évolution est donc programmée comme l’est le développement de l’embryon.
Pour montrer cela, il faudrait montrer que l’évolution continue dans le même sens même lorsque l’environnement a été totalement perturbé, ou que la même évolution se fait dans des lignées cousines soumises à des environnements différents. Il s’agit alors d’une orthogenèse. Lamarck propose ainsi une orthogenèse lorsqu’il inscrit une tendance au progrès dans le vivant qui fait évoluer les êtres vivants des vers les plus simples jusqu’à l’Homme, l’effet de l’usage et du non usage, lié à l’environnement, n’étant qu’une force secondaire qui vient perturber et dévier un peu l’orthogenèse.
Dans son ouvrage Structure de la théorie de l’évolution Stephen Jay Gould plaide pour réconcilier sélection et déterminisme interne. Il y a des contraintes liées au développement et au fonctionnement. La sélection n’est pas ici toute puissante et la variation n’est pas isotrope. On pourra parler de canalisation.
Cela concilie aussi les deux pôles structuraliste et fonctionnaliste. Il existe des plans d’organisation communs qui sont un héritage reçu d’un ancêtre. Les êtres vivants dans leur évolution doivent composer avec cet héritage, qui est à la fois une restriction, car il impose certaines limites, mais aussi une canalisation, c’est-à-dire qu’il ouvre un champ de possibles dans certaines directions.
L’adaptation à une fonction se surimpose ensuite, modulant le plan de base. Par rapport à la variation isotrope de Darwin, on donne plus de poids aux contraintes.
Mais canalisation n’est pas tendance : si la variation est canalisée, elle n’est pas pour autant orientée vers une finalité inscrite on ne sait où. Les contraintes internes peuvent faire qu’une direction se trouve privilégiée sans que cela corresponde à un plan inscrit.
Dans le prolongement de ces questions de structure se trouve celle sur le rôle des organes en cours d’évolution : à quoi sert une demie-aile ?
La réponse apportée par les évolutionnistes a été celle de l’exaptation : un organe qui joue un certain rôle se retrouve récupéré pour remplir une autre fonction. On retrouve là le bricolage de l’évolution.
L’étude d’une structure peut finalement s’envisager sous au moins trois angles pour rendre compte de son existence et de ses caractéristiques :
-
-
- Fonctionnaliste, c’est-à-dire sous l’angle de l’adaptation à la fonction, façonnée par la sélection naturelle. C’est l’angle qui a prévalu pendant une large part du 20ème siècle et qui a conduit aux excès du pansélectionnisme. Pour éviter cet écueil, il faut garder à l’esprit les deux autres angles.
-
-
-
- Historique et phylogénétique : la structure présente est héritée et liée à l’histoire de la lignée,
-
-
-
- Structuraliste, c’est-à-dire sous l’angle des contraintes de structure, de construction et de fonctionnement de l’organisme.
-
- Une orientation de la variation par l’environnement ?
Dans la théorie de l’évolution se répercutent les échos de vieux débats : structuralisme vs fonctionnalisme comme on l’a évoqué par exemple. Une autre question épineuse s’y retrouve, celle de l’inné contre l’acquis, doublée par celle de l’interne contre l’externe.
Lamarck comme Darwin utilisaient dans leurs théories une hérédité des caractères acquis : sous l’influence de l’environnement, l’individu se transforme, acquiert un nouveau caractère qu’il pourra transmettre à sa descendance. Cette hérédité a été disqualifiée par la séparation germen/soma définie par Weismann et par les découvertes des bases de l’hérédité.
Pourtant cette notion d’hérédité de caractères acquis opère un retour partiel en grâce grâce à des découvertes récentes, mais sous une forme différente.
Les études sur la réalisation des caractères ont montré que l’environnement pouvait intervenir à plusieurs niveaux. Il peut agir sur le produit du gène, c’est-à-dire sur la protéine, qui par ses propriétés ou son activité détermine le caractère. Il peut aussi agir au niveau des gènes, non pas en les modifiant mais en modulant leur expression, c’est-à-dire que les gènes s’exprimeront (la protéine sera synthétisée) en fonction des conditions expérimentales. On voit que la conception est différente de ce que pensaient les biologistes à l’époque de Lamark ou de Darwin car ici l’environnement ne modifie pas l’information génétique. Il ne peut faire s’exprimer que ce qui est déjà écrit.
Cette sensibilité à l’environnement est variable selon l’espèce considérée mais elle existe. Le phénotype est plastique : il peut se modifier en réponse à l’environnement. Plusieurs phénotypes sont possibles à partir d’un même génotype.
Or on s’est aperçu que certains de ces phénotypes modifiés par l’environnement étaient transmissibles à la descendance, et que les caractères pouvaient rester même lorsque le facteur environnemental qui les a produit n’est plus présent.
Plus qu’une transmission de l’information génétique lors de la reproduction, il peut aussi y avoir une transmission de l’état d’expression de cette information. On parle d’hérédité épigénétique. (Concrètement, cela correspond par exemple à l’état de méthylation de l’ADN, qui va pouvoir inactiver certains gènes.)
Pour en savoir plus : le Darwinisme évolue aussi par Eva Jablonka, dossier La Recherche mai 2007 L’évolution
- Un rôle des virus
Une proposition récente est un rôle possible des virus dans l’évolution du monde vivant, dans l’évolution des génomes. Certaines caractéristiques et certaines fonctions auraient été apportées par des virus. L’ADN et le noyau seraient une invention virale. La syncitine, une protéine essentielle dans la formation du placenta, serait en fait une protéine de rétrovirus, intégrée dans le génome de l’organisme et recrutée par les Mammifères, qui profitent ainsi des techniques développées par les virus pour échapper au système immunitaire pour que le fœtus ne soit pas reconnu comme étranger et rejeté par l’organisme maternel.
Pour en savoir plus :
– Une conférence de l’UTLS : Nouvelles questions et hypothèses sur l’origine et l’évolution des génomes de Patrick Forterre
– Des virus ont-ils inventé l’ADN ? par Patrick Forterre, dossier Pour La Science Où est née la vie ?
Cela comme les familles multigéniques illustre le bricolage à l’œuvre dans l’évolution, pour reprendre le mot de F. Jacob dans le Jeu des Possibles.. Il n’apparait que peu de choses nouvelles et il s’agit davantage de réutiliser ce qui existe déjà.
- Le tempo de l’évolution : La théorie des équilibres ponctués
Pour Darwin, l’évolution procède sur un mode gradualiste : les changements sont insensibles et ne deviennent perceptibles qu’au bout d’une longue durée. Ils se font par l’accumulation de petites variations. Darwin réinvestit ici les notions introduites par Charles Lyell, qui avait posé les bases de la géologie en définissant ce mode d’accumulation insensible pour expliquer les phénomènes géologiques. Cela correspond à l’uniformitarisme par opposition au catastrophisme, qui utilisait des cataclysmes dans ses explications. Charles Darwin n’a eu de cesse d’appliquer cet uniformitarisme, depuis son étude sur les îles océaniques jusqu’à celle sur l’action des Lombrics.
Or dans les archives géologiques, on ne voit pas ces changements graduels mais on observe une relative stabilité des formes jusqu’à certains niveau où apparaissent brusquement de nouvelles formes.
L’évolution ne semble pas avancer si tranquillement. Darwin se reposait sur l’incomplétude des archives fossiles pour répondre à cette contradiction entre sa théorie et les observations.
Peu satisfaits, Gould et Eldredge proposèrent leur théorie des équilibres ponctués, qui retrouve un certain goût de catastrophisme. L’histoire des espèces est faite de longues stases pendant lesquelles les formes changent peu et de brusques ponctuations pendant lesquelles les formes vont diverger rapidementsous l’effet d’un changement majeur de l’environnement. On quitte ici le mode gradualiste pour se rapprocher d’un saltationnisme : évolution par sauts.
L’élucidation de la disparition des Dinosaures (et de nombre de leur contemporains) sous l‘effet de l’impact d’une météorite il y a 65 Ma va dans ce sens.
On a pu aussi mettre en évidence qu’un environnement perturbé engendre plus de mutations chez les Bactéries et de recombinaisons chez certains végétaux. Tout se passe donc comme si certains systèmes de correction étaient levés pour l’occasion, pour augmenter la probabilité de produire une variation avantageuse.
Pour en savoir plus : Mutation et nouveauté par Jacqueline Laurent, bibliothèque Pour La Science : L’évolution
Le développement embryonnaire est normalement assez peu sensible aux effets de l’environnement, grâce à certains systèmes de sécurité comme les protéines chaperonnes qui aident les autres protéines à se replier sur elles mêmes et vont pouvoir ainsi masquer l’effet de certaines mutations. Lorsque les conditions environnementales varient beaucoup, ces systèmes de sécurité pourraient sauter et le développement devient alors plus sensible à l’environnement, produisant des formes plus variées sur lesquelles la sélection naturelle peut agir. On assiste alors à une accélération de l’évolution, avec la production de plus de variants.
Pour en savoir plus : comment le milieu sculpte les gènes par Vincent Débat, Dossier La Recherche mai 2007 L’évolution
La stase ne signifie pas qu’il ne se passe rien, que l’évolution est nulle. Des changements mineurs, internes, invisibles sur les fossiles, se produisent. Les fréquences allèliques varient. Mais la forme générale peut n’être que peu modifiée. C’est ce qu’il se passe pour le Coelacanthe, poisson dont la forme semble être la même depuis des millions d’années.
Pour en savoir plus : Les fossiles vivants n’existent pas - Armand de Ricqlès, bibliothèque Pour La Science : L’évolution
L’évolution est nécessairement permanente. Leigh Van Valen a montré que quelle que soit la durée d’existence déjà écoulée, la probabilité d’extinction reste la même. Autrement dit quel que soit le temps passé à évoluer on n’est jamais parfaitement adapté à l’environnement. Evoluer est un travail constant. Car l’environnement n’est jamais constant : même si l’environnement physique reste le même, il y a dans l’environnement de l’individu d’autres individus, d’autres espèces, avec lesquelles il va interagir, positivement (coopération comme dans le cas des plantes à fleurs et des insectes) ou plus négativement (prédation, parasitisme par exemple). Tout le monde évolue donc sous l’effet de ces interactions, chacun doit évoluer en réponse à l’évolution des autres. On parle de coévolution. Van Valen a nommé cela hypothèse de la Reine Rouge, d’après un passage de De l’autre côté du miroir de Lewis Caroll : il faut courir vite pour pouvoir rester sur place. Il faut continuer à évoluer pour pouvoir rester.
Pour en savoir plus : Leign Van Valen et l’hypothèse de la Reine rouge - Claude Combes, bibliothèque Pour La Science : L’évolution
Pour en savoir plus :
– Une conférence de l’UTLS : Combien de temps faut-il pour faire une espèce ? de François Bonhomme
- Quels niveaux de sélection ?
Pour Darwin, la sélection opère au niveau de l’organisme. Richard Dawkins a voulu avec son gène égoïste ramener cet organisme à un vecteur de gènes, la sélection opérant au niveau de ces gènes. Gould plaide lui pour étendre la sélection à tous les niveaux, depuis le gène jusqu’aux clades, avec éventuellement différents degrés, de façon à mieux comprendre aussi la macroévolution.
- Passage à la macroévolution
Nous restons ici dans des questions d’échelle et de rythme.
Ces termes de microévolution et macroévolution ne sont pas reconnus par tous les évolutionnistes, en partie parce que cette distinction entre les niveaux d’évolution a été introduite par des non darwiniens tenants d’un saltationnisme et d’une orthogenèse, qui en quelque sorte voulaient bien reconnaître la sélection naturelle au sein de l’espèce mais pas au-delà (ce qui est une façon de limiter la portée des idées de Darwin).
Dans la théorie synthétique, le couple variation/sélection à l’échelle de l’organisme suffit à expliquer ce qu’il se passe à l’échelle de l’espèce ou du clade par extrapolation, avec une durée de temps assez longue, ainsi que le proposait Darwin. La macroévolution est la somme de microévolutions.
Gould propose dans une théorie hiérarchique de l’évolution de distinguer ces différents niveaux, dans un mouvement opposé à celui de Richard Dawkins qui ramène la sélection au seul niveau du gène. Gould replace à d’autres niveaux les mécanismes décrits au niveau de l’individu, éventuellement avec des poids différents selon le niveau, tout ceci cadrant avec sa théorie des équilibres ponctués.
La découverte des gènes du développement, notamment avec l’étude de la Drosophile et de ses mutants, a ouvert de nouvelles perspectives. L’existence de ces gènes architectes, qui commandent la construction de l’organisme pendant l’embryogenèse, et l’observation des modifications produites par leur mutation a laissé voir que dans certains cas une mutation sur un seul gène peut avoir des conséquences multiples et importantes, du fait de la position de contrôle qu’exercent ces gènes du développement, et donc modifier le plan d’organisation d’un organisme.
On voit ici une possibilité de saltationnisme, dans le cadre d’une macroévolution, qui rappelle le mutationnisme de de Vries ou la théorie des monstres prometteurs de Goldschmidt. La polyploïdie était un autre type de macromutation susceptible de faire apparaître brusquement de nouvelles espèces.
Cela a conduit à un regain d’intérêt pour l’étude du développement embryonnaire, couplée à l’étude de l’évolution, dans une approche appelée évo-devo, où la génétique tient une place importante. Le premier résultat important de cette approche à été la surprenante découverte de la conservation des gènes du développement dans tout le monde animal. Un autre résultat a été l’importance de facteurs cytoplasmiques, mis en place par la mère, dans les premières étapes du développement, avant que les gènes ne prennent le relais. La comparaison des développements de diverses lignées montre comment des modifications dans les étapes de développement permettent de comprendre les différences entre plans d’organisation.
Pour en savoir plus :
deux conférences en ligne de la cité des sciences :
– Les gènes du développement, architectes de l’embryon
– L’évo-dévo en débat
– Les gènes du développement par William McGinnis et Michael Kuziora, bibliothèque Pour La Science : L’évolution
– Les mutations des gènes Hox chez les Mammifères par Pascal Dollé,bibliothèque Pour La Science : L’évolution
La théorie de l’évolution est encore riche de questionnements et de pistes de recherches. Il manque aujourd’hui une nouvelle synthèse pour intégrer les découvertes récentes. Cette prochaine synthèse, à l’image de la théorie hiérarchique de Stephen Jay Gould, devra concilier plus que trancher entre les différentes conceptions (gradualiste ou saltationniste, fonctionnaliste ou structuraliste, échelle du gène, de l’organisme ou de l’espèce, importances respectives du hasard et de la sélection...), tant il est vrai que dans le monde vivant, foisonnant et complexe, il y a rarement une réponse simple et unique.
Cette complexité de la théorie, qui est en réalité davantage un agrégat de microthéories autour de la thèse centrale du couple darwinien variation / sélection, est un premier obstacle à une bonne réception de la théorie de l’évolution.
– En premier lieu, cette complexité rend très l’évolution très difficile à se représenter, à cause du mode gradualiste, de la part de hasard, de l’échelle de la population, des temps très longs impliqués.
Un argument classique à l’encontre de l’évolution est la question de l’utilité d’un organe pas finalisé, un demi-œil ou une demi-aile. C’est ce que Richard Dawkins appelle l’Argument d’Incrédulité Personnelle. On note au passage qu’il est présupposé qu’il faut absolument que la structure serve à quelque chose. Nous sommes dans une représentation de la Nature parfaite, optimisée, finalisée.
L’exaptation, c’est-à-dire l’acquisition d’une nouvelle fonction par une structure préexistante qui remplissait jusque là une autre fonction, est un élément de réponse. L’exemple classique est la plume, qu’on pense d’abord impliquée dans la thermorégulation et qui s’est trouvée favoriser le vol.
Un autre élément de réponse est que même la structure non finalisée dans son état actuel (d’ailleurs pas nécessairement parfait) peut quand même avoir une utilité. Une couche de cellules capables de capter la lumière est déjà utile pour distinguer le jour de la nuit, percevoir l’ombre d’un prédateur qui passe. Il n’est pas nécessaire d’avoir un œil avec un système de mise au point amélioré et une bonne acuité visuelle pour qu’il soit utile.
On pourra donc montrer à qui donne ce premier argument contre le mécanisme de l’évolution que ce n’est pas parce qu’il est difficile d’imaginer à quoi sert un demi-œil que ce demi-œil ne sert à rien.
– En second lieu une trop grande simplification, l’utilisation de mauvais termes ou de certains exemples va créer une représentation fausse de l’évolution, sur laquelle vont s’articuler des rejets de la théorie pour des raisons extérieures à la science. Nous les enseignants avons ici une grande part de responsabilité.
Ainsi nous n’insistons pas suffisamment sur le fait que l’évolution ne se fait pas tant sur une survie différentielle grâce à certains caractères que par un accès différentiel à la reproduction, ce qui n’est pas la même chose. Parle-t-on assez de reproduction plutôt que de survie avec l’exemple de la Phalène du Bouleau ?
Ainsi dans les images classiques des documentaires animaliers sur des troupes de grands Mammifères, la sélection ne se fait pas tant au travers de la mort sous les crocs des prédateurs, qui éliminent d’abord les plus vieux et les plus malades parce que ce sont les plus faciles à attraper, qu’au travers des combats entre mâles pour les femelles, combats qui se terminent rarement par la mort du vaincu. Or l’expression classique de lutte pour la vie (struggle for life) dépeint une évolution basée sur la survie des plus forts aux dépends des plus faibles. Voilà une image qu’il nous faut tempérer, parce qu’elle est fausse, et parce qu’en plus elle génère une image de la Nature qui peut conduire à un rejet de la théorie de l’évolution pour des raisons morales. C’est un argument classique des opposants à la théorie de l’évolution, qui l’accusent d’être à l’origine de bien des maux de l’histoire récente.
On s’apercevra que beaucoup d’obstacles moraux ou métaphysiques à la réception de la théorie de l’évolution sont en fait basés non pas sur la réalité de la théorie mais sur des caricatures, sur une mauvaise compréhension de la théorie.
II. Hasard et finalisme
III. Vitalisme et mécanisme
IV. La place de la Science, Religion et Morale
V. Enseigner l’évolution de la lignée humaine
Me contacter : B. Boucher
Une réaction, une correction ? Voir le forum à la suite de cet article (Répondre à cet article).
Bibliographie.
– L’évolution, sous la direction d’Hervé Le Guyader, Belin Pour La Science
– Dossier La Recherche mai 2007 L’évolution
– Dossier Pour La Science juillet 2008 Où est née la Vie ?
– L’origine des espèces - Charles Darwin, GF Flammarion
– Structure de la théorie de l’évolution - Stephen Jay Gould, NRF Essais
– La Vie est belle - Stephen Jay Gould, Seuil
– Evolution, synthèse des faits et théories - Francine Brondex, ed DUNOD
– Les avatars du gène - Pierre Henri Gouyon, Jean Pierre Henry, Jacques Arnould, Belin Pour La Science
– Le fleuve de la vie - Richard Dawkins, ed Hachette